A.Soosaar ¤¤ Märkmeid füsioloogiast 2022–
Peritsüüdid
Aastakümnete jooksul olen kuulnud kümnetest erinevatest tsüütidest või blastidest ning õppinud neid ka rohkem või vähem tundma, kuid alles nädalapäevad tagasi jõudis ühe JoP artikli kaudu minu teadvusse tõsisem teadmine peritsüütide olemasolust ja rollist organismis. Võimalik, et olin nendest midagi kuulnud juba ka varem (n.t kunagi 1980. aastate teisel poolel, kui tegin farmakoloogia kateedri seminaril ülevaate hematoentsefaalsest barjäärist), kuid varasemast siiani säilinud mälestust nende olemasolu, rolli ja tähenduse kohta pole ometi tekkinud. Lihtsalt termini enda etümoloogiast lähtuvalt võiks tegemist olla millegi ümber või midagi ümbritsevate rakkudega ja teaduse abiga saabki selgeks nende paiknemine eeskätt kapillaaride või täpsemalt nende endoteeli rakkude ümber. Olulisemad inimese füsioloogia õpikud saavad tänase päevani peritsüütideta hakkama, vaid Boroni ja Boulpapi põhjalikus õpikus mainitakse neid mõnel korral hematoentsefaalset barjääri ja mikrotsirkulatoorset ala loova struktuuri selgitamisel, kuid funktsiooni ometi täpsemalt kirjeldamata. Tõele au andes on füsioloogia õpikud kuni eelmise sajandi lõpuni õige kidakeelsed ka endoteeli rakkude funktsionaalse rolli ja seda tagavate protsesside süsteemsel kirjeldamisel. Pigem peeti kapillaaride seina õige passiivseks ja staatiliseks barjääriks vere ja koevedeliku vahel, mille omadused ajas ei muutu ning ainete liikumine läbi kapillaari seina on määratud pigem mõlemal pool oleva keskkonna omadustest (n.t enamus Starlingi valemi komponentidest ehk jõududest). Ju oli neil aegadel füsioloogia võitlusväljaks ikka suuremate mõõtmetega objektid ja süsteemid ning neis ilmnevad efektid, aga nendes toimiv tegelik aineline kooslus jäi vahendilise suutmatuse tõttu sageli üpriski mõistmatuks. Peritsüütide kohta käiva teadusliku uudishimu seisu annab päris hästi Pubmedi vastav teemakohaste viidete/kirjatööde arvu graafik, millest näeme, et eelmise sajandi 1975. aastani ilmus peritsüütide kohta mõni artikkel aastas, ja varasematel aastatel mõnikord üldse mitte, ning sealt edasi hakkab aastas ilmuvate artiklite arv järjest kiiremini kasvama ning hoog läheb päris suureks sel sajandil, saavutades viimastel aastatel taseme üle 500 teadustöö aastas. Siinkohal tasub meenutamist, et esimesed kirjalikud teated peritsüütide olemasolu kohta pärinevad 1870. aastate algusest ning nende nimi pärineb alles 1923. aastal avaldatud Karl Wilhelm Zimmermanni kapillaaride ehitust kirjeldavast tööst.
Põgus tuur internetis andis aimu, et peritsüüdid on tänapäeva rakubioloogias ja sellest lähtuvates füsioloogia ja kliinilise meditsiini valdkondades järjest olulisemad tegijad nii normi kui haiguse puhul. Lühikese ringi luusimise käigus oli seal hea soe tunne kohata Eesti päritolu teadlast Annika Kellerit, née Armulik, kes olnud varasemalt selle sajandi heade ülevaadete esimeseks autoriks ning värskemalt juba nende rakkude rolli konkreetsemate aspektide oluliseks uurijaks. Ei hakkakski rohkem dr Guuglit otsimise palvega tülitama ja piirdun järgnevas dr A. Kelleri osalusel 2005. ja 2011. aastal avaldatud ülevaatliku kirjasõnaga, sest neis sisaldub päris palju sellest teadmisest, mis praeguseks peritsüütide kohta üldse olemas on.
Kui eelmise sajandi alguse tööd keskendusid eeskätt peritsüütide väljanägemisele nähtuna valgusmikroskoobis, siis hiljem on seda pilti täiendatud andmetega järjest suurema „nägemisevõimega“ vaatlusriistadest, n.t elektronmikroskoobist. Erinevad ülevaated toovad „parema nägemisvõimega“ riistadelt saadud piltide põhisisu kokkuvõtlikult välja, näiteks on täpsemalt selgeks tehtud mõlemat tüübi rakkude ja basaalmembraani omavahelise füüsilise kontakti viisid, millest omakorda mulkühendused võimaldavad ainete ja infovahetust erinevate rakkude vahel. See basaalmembraan on ka üks põnev koht, mille tekkest, omadustest ja talitlusest füsioloogias eriti juttu ei tehta. Samuti on tuvastatud peritsüütide väljanägemise mitmekesisus, erinev arvukus erinevates kudedes (lihaste kapillaaride ümber on neid 100 korda vähem kui endoteliotsüüte ning aju ja silma kapillaarides on nende arv enam-vähem võrdne) ning teatud omaduste sarnasus nende lähedal paiknevate silelihasrakkudega. Samas näikse peritsüüdid olema ikkagi jätketega rakud, mis kontakteeruvad mitme endoteliotsüüdiga (vt joonis 1).

Joonis 1. Skemaatiliselt esitatud mitut tüüpi peritsüütide, silelihasrakkude ja endoteliotsüütide väljanägemine ja paiknemine üksteise suhtes.
Allikas: Microvascular bioengineering: a focus on pericytes | Journal of Biological Engineering | Full Text (biomedcentral.com)
Armuliku ja kaasautorite 2005. aasta ülevaateartikkel keskendub peritsüütide suhtlusele endoteliotsüütidega. See kommunikatiivne partnerlus võiks ju üksikute rakkude tasemelt vaadatuna mõneti veidralt kõlada, kuid ülevaate suureks tõdemuseks on arusaam, et mikrotsirkulatoorse ala (rakkudest juba vähemalt üks tase ülespoole või keerukamale) normaalse toimimise aluseks on hädatarvilik teatud interaktsioon peritsüütide ja endoteliotsüütide vahel või teisipidi seda väljendades, puuduliku interaktsiooni puhul häirub kogu piirkonna funktsioneerimine. Taaskord on ju tegemist konkreetse juhtumiga struktuuri-funktsiooni kooskõla põhimõtte rakendumisel, mille abil füsioloogias sageli seletatakse nii normaalset kui haiguslikku talitlust. Robustne mõttelõng võiks siin olla selline, et teatud haiguslike protsesside tulemusena väheneb kahjustunud kapillaaride ümber peritsüütide hulk või kaovad nad sootuks ning selle tulemusena häirub interaktsioon kahe rakutüübi vahel, mis omakorda häirib moel või teisel ainete vahetamist vere ja kudede vahel ja tulemuseks koe spetsiifilisemate funktsioonide häire. Kõnealuses töös keskendutakse nimetatud interaktsiooni käsitlususes keemilisele rakusignalisatsioonile, mida iseloomustatakse konkreetsete virgatsainete ning nendega seotud info ülekandemehhanismide alusel. See rakusignalisatsioonil põhinev suundumus on füsioloogias viimaste aastakümnetega saanud universaalseks ning selle ajendiks on uurimismeetodite loomine, millega on võimalik rakus/organismis olevaid aineid ja nende omadusi ühe täpsemini uurida. Ülevaatest selgub, et 1990. ja 2000. aastate alguses tehakse kindlaks transformeeriva kasvufaktori beeta vormi (TGFβ), angiopoetiini, vereliistakutest pärit kasvufaktori informatsiooniline toimetamine peritsüütidel ning suhtluses osalevate ainete arv sealt edasi on järjest kasvanud.
Kõrvalepõikena olgu märgitud, et umbes saja aastaga on arusaam rakusignalisatsiooni mitmekesisusest märkimisväärselt avardunud. Kõigepealt tehti 20. sajandi esimestel kümnenditel kindlaks mõnede hormoonide (sekretiin, insuliin, suguhormoonid) ja neuromediaatorite (atsetüükoliin ja noradrenaliin) olemasolu ja püüti üksnes nendega seletada organismis toimuva reguleerimist. Küllap oli tookordetes uut moodi seletustes ikkagi hulgaliselt raskusi organismis toimuva kirjeldamisega, mis omakorda innustasid uurijaid infoülekandega organismi sees edasi tegelema. Nii ongi tänaseks päevaks teadmistega jõutud välja arusaama, kus konkreetsete rakkude vahelises suhtluses osaleb kümneid kui mitte sadu erinevaid signaalaineid, nende retseptoreid ja sihtrakus olevaid signaaliradu (NB! Keegi ilmselt ei tea täpset nende koguarvu tervikorganismis), mis omakorda täidavad elusa sisemises suhtlusprotsessis erinevaid rolle alates sõnumi moodustumisest ja kandmisest kuni selle ära tundmise ja sisuks oleva sõnumi mõju realiseerumiseni. Teiseks toovad selle artikli autorid esile kõnealuse interaktsiooni uurimise metoodika, mis põhineb erineval viisil geneetiliselt muundatud organismides toimuva uurimisel. Geneetilise muundamise sisuks on kõigepealt signaalainetega seotud valke determineerivate geenide väljalülitamine, üleaktiveerimine jms ning seejärel tehtud muundamise mõju uurimine. Seegi on äsja mainitud struktuuri-funktsiooni ühtsuse (või isegi ühtsuse kindlakujulise rikkumise) põhimõtte kohaldamine üksikute ainete ja nende vastasmõju tasemele. Füsioloogia ja närviteaduse arengus on selle põhimõtte erinevatel organisatsiooni tasemetel eksperimentaalne rakendamine olnud väga levinud ning andnud hulgaliselt uut olulist teadmist organismi ja selle osade talitlemise kohta. Nii on aju erinevate osade funktsioone esimeses lähenduses uuritud just nõnda, et katseloomadel purustatakse üks või teine struktuur ja selgitatakse selle tagajärjed või seostatakse patsientidel ilmnev kliiniline pilt lahangul leitud struktuurse muutusega.
Kui vaadata lähemalt Armuliku ja kaasautorite 2011. aasta ülevaadet, siis selle haare on oluliselt laienenud ning eesmärgiks juba peritsüütidest terviklikuma pildi esitamine. Samas mõneti üllatuslikult toovad autorid välja raskused peritsüütide ühemõttelise identifitseerimise ja defineerimisega, sest polevat täiesti spetsiifilisi peritsüütide tunnuseid. Küllap identifitseeritaksegi peritsüüte nii morfoloogiliste kui biokeemiliste tunnuste kombinatsioonina ning mure on mingi spetsiifilise markeri või tuntud markerite spetsiifilise kombinatsiooni tuvastamisega. Ortodoksses funktsionalismis ei peaks toimetava üksuse päritolu olema primaarse tähendusega, peamine on ikka toimetamiste mõistmine ise. Samas ei ole teema päris tähtsuseta, sest mitmelt tuleb tõendusi peritsüütide heterogeensuse kohta, mis edaspidi võib ikkagi jõuda punkti, praeguses arusaamas peritsüütide hulka hoopiski erinevat tüüpi ja erineva võimekusega rakud. Nii näikse ka ülevaate autoritel olema veendumus, et peritsüüdid ei kujuta endast homogeenset rakkude rühma, vaid erinevate kriteeriumide alusel saab nende seas eristada veel alarühmigi (midagi sellist on näha ka joonisel 1). Autorite lemmikteema on jätkuvalt infot vahetavate ainetega seonduv peritsüütide kandis. Minemata detailidesse peaks nende artikli vastav joonis (meie loos joonis nr 2) andma tunnetuse peritsüütidega seotud sõnumivahetuse paabelist, mis kindlasti ei ole veel kogu tõde selles suhtluses osalevatest keeltest, süsteemide keemilise kineetika funktsionaalsed võimalused ei ole siin veel üldse esitatud.
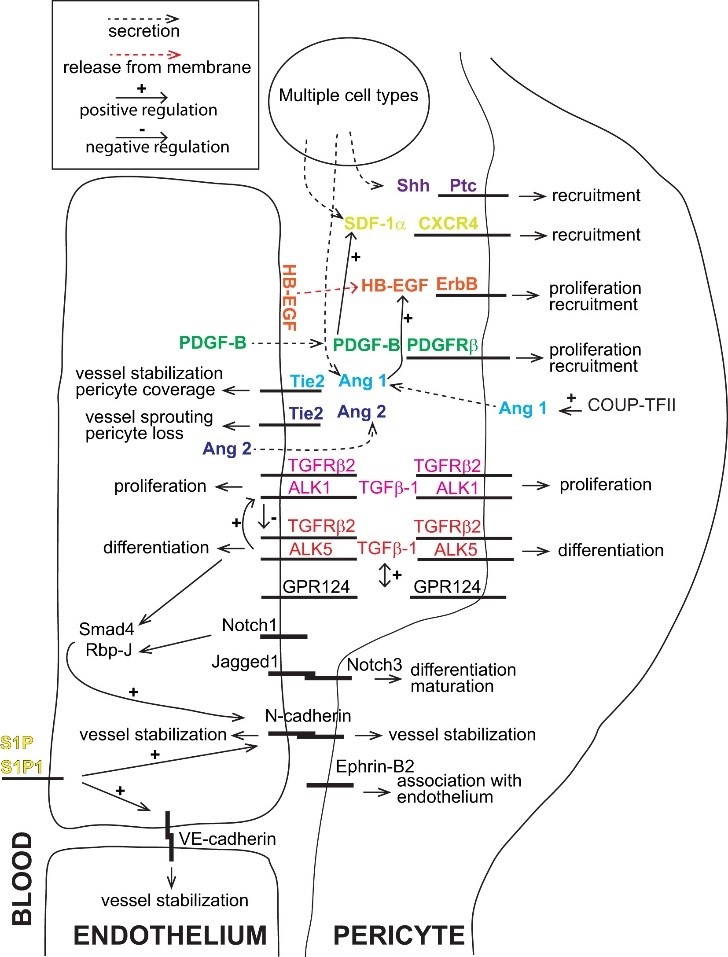
Joonis 2. Armulik et al. (2011) artiklist pärit graafiline ülevaade keemilisest sõnumivahetusest peritüütide ja endoteliotsüütide vahel. Täpsemaks vaatluseks klikka pildile.
Allikas: https://doi.org/10.1016/j.devcel.2011.07.001
Kui võtta nüüd lähemale vaatlusele ülevaates esitatud peritsüütide funktsioonide kirjeldamine, siis sealt nähtub peamise makrotõdemusena, et peritsüütide puudumine või siis ka defitsiit toob kaasa kapillaaride läbilaskvuse suurenemise, millest järeldub kiiresti, peritsüüdid osalevad selle määratlemises, mis üldse läbi kapillaaride seina liikuda saab. Muidugi saab makronähtusega sobitada ka hematoentsefaalse barjääri olemasolu, sest eks ole seegi ajus ühe ainetele selektiivse transpordi barjääri loomine. Ja kui napib korralikult toimivaid peritsüüte, ei tööta ka barjäärid ainete liikumisele vajalikul moel. Mitte kuigi selge on peritsüütide rolli pisikeste veresoonte diameetri regulatsioonis. Arvestades kontraktiilsete valkude olemasolu ja talitlust peritsüütides ja signaalainete rohkust piirkonnas ei sugugi võimatu, et saab nii otseselt kui kaudselt silelihasrakkude mõjutamise kaudu peritsüütidest pärit ainetega soonte diameetrit reguleerida. Suur lootusi pannakse peritsüütide rollile angiogeneesis mis on omakorda tähtis nii kasvu kui regeneratsiooni korraldamisel. Kõigi kolme funktsiooni puhul näikse seletused ühel hetkel jõudvat punkti, kus keemilise faktori mõju konverteerub ümbruskonnale avaldatava mehhaanilise mõju muutuseks, mis võib siis väljenduda soonefiltri läbilaskvusomaduste või soone diameetri muutmises või hoopis soone seina elementide juhatamises kuhugile uude kohta. Ei tundu kuigi usutav olevat, et signaalained ise seda kõike suudavad, küllap tulevad ikkagi mängu nende abil täpselt juhitud rakumehhaanika protsessid.
Jah, tõesti üllatav, kui vähe on kuni viimase ajani peritsüütide talitlusest juttu olnud ning kui suurt ja olulist tulevikku praeguse uuringud neile omistavad. Samas jääb neile tänase päeva arusaamade järgi ikkagi pigem endoteeli rakke täiendav või abistav roll, mitte suuresti iseseisev ja seni tundmatu laialdase mõjuga struktuuri-talitluse seose ilming. 5
- oktoober 2022